Que se passe-t-il dans le cerveau d'un bébé à la naissance ?

La naissance d’un enfant est un moment extraordinaire dans la vie d’un parent. Assister à la naissance de son enfant procure des émotions que nulle autre situation ne peut provoquer. Mais cette situation est encore plus spéciale pour le nouveau-né, qui doit faire preuve d’une sacrée capacité d’adaptation ! L’espace de quelques minutes, il doit effectuer ce que l’évolution a mis plusieurs millions d’années à faire : s’adapter de la vie sous-marine à la vie terrestre !
La naissance est ainsi une période de transition majeure pour le nouveau né. Ils quittent leur cocon, stérile et parfaitement régulé à 37°C, pour un environnement hostile et froid ! L’oxygène nécessaire à sa survie ne provient plus du placenta mais de ses poumons, nécessitant leur bon déploiement et une bonne vascularisation sanguine. La naissance correspond donc, du point de vue du bébé, à une période de stress intense !
Comment le cerveau du bébé arrive-t-il à s’adapter à ce moment si particulier ? Comment fonctionne-t-il dans le ventre de sa mère, et comment doit-il s’adapter à l’environnement extérieur ? Comment les hormones maternelles protègent-elles le cerveau fœtal ?
Il faut savoir être flexible ! [1]
L’Homme possède deux caractéristiques marquantes quand on le compare à ses compères primates : il possède un gros cerveau et marche sur ses deux jambes. Cela est bien pratique dans la vie quotidienne, mais nous allons le voir, cela est bien plus problématique lors d'un accouchement. La bipédie nécessitant un bassin plus étroit, elle engendre fatalement un espace restreint lorsque le bébé doit quitter le ventre de sa mère ! La taille de la tête (qui est le facteur limitant du côté du bébé) et du bassin maternel doivent donc être parfaitement adaptés. Si ce n’est pas le cas (si le bébé est « trop gros » ou le bassin de la maman « trop étroit »), on pourra avoir recours à la césarienne pour une naissance en toute sécurité.
Les deux partenaires doivent donc faire preuve de flexibilité au cours de la naissance… au sens premier du terme ! En effet, les os du bassin maternel se déplacent légèrement pour maximiser l’ouverture de la filière génitale. Le corps du bébé, et en particulier son crâne, est soumis à rude épreuve.
C’est ce qu’a mis en évidence une drôle d’étude, française, publiée en 2019 [1]. Afin de mettre en évidence les déformations du crâne et du cerveau du nouveau-né au cours de la naissance, ils font fait passer une IRM à des femmes… en plein travail. Tentez d’imaginer ces femmes dans une salle d’imagerie en train de passer une IRM alors même que leur enfant est en chemin !
Fort heureusement, l’IRM se trouvait juste à côté de la salle de naissance et la procédure d’acquisition des images (qui peut être bien longue d’habitude!) avait été optimisée pour durer au plus une douzaine de minutes.
Les scientifiques mirent en évidence une déformation (normale!) importante du crâne et du cerveau du bébé, qui n’avait été que modélisé jusqu’alors. Les différents os constituants le crâne (les os frontaux, pariétaux, occipitaux ou encore l’os sphénoïde) se superposent, voire se chevauchent à certains endroits, comprimant le cerveau qui se déforme lui aussi ! La pression subie lors du passage de la filière génitale aboutie à une déformation dite « en pain de sucre », qui est heureusement la plupart du temps rapidement réversible une fois le bébé sorti !
 |
| Le Pain de Sucre de Rio de Janeiro (pour vos donner une idée de la déformation du crâne de bébé ! Le téléphérique en moins). |
Le cerveau d’un bébé est donc soumis à d’importantes contraintes et à un stress biologique majeur. Au delà des contraintes physiques, comment évolue-t-il au cours de ces quelques heures cruciales ? Comment se protège-t-il des dangers et des multiples agressions ? Nous allons le voir, cela implique des changements majeurs dans le fonctionnement des neurones !
Comment fonctionne un neurone ? [6, 7, 8, 9]
Un neurone, comme toute cellule du corps humain, est limité par une membrane lipidique. Cette membrane, normalement imperméable aux ions et aux molécules d’eau, sépare de façon hermétique le milieu intra- du milieu extra-cellulaire. La composition de ces deux milieux en ions est différente : la concentration en sodium (Na+) est plus importante dans le milieu extra-cellulaire (140mmol/L versus 20mmol/L) et celle en potassium (K+) est plus importante dans le milieu intra-cellulaire (150mmol/L versus 4mmol/L).
Ces concentrations, activement maintenues par des protéines membranaires, aboutissent à un potentiel membranaire, égal à -70mV. Ainsi donc, l’intérieur de la cellule est chargé négativement, alors que le milieu extracellulaire est chargé positivement.
 |
| Concentrations en ions dans et autour d'un neurone. |
Il existe donc, de part et d’autre de la membrane cellulaire, un gradient à la fois chimique (c’est à dire de concentration en ions) et électrique (une différence de potentiel). Ces gradients sont maintenus par l’imperméabilité et l’isolation électrique de la membrane cellulaire, ainsi qu’au fonctionnement continu de protéines membranaires (notamment la pompe ionique Na/K ATPase) dont le rôle est de chasser le sodium hors de la cellule et de pomper le potassium du milieu extra-cellulaire.
Ces gradients chimiques et électriques ne sont pas indépendants l’un de l’autre. Concernant le sodium, le gradient de concentration le pousse à rentrer dans la cellule pour équilibrer les concentrations intra et extra-cellulaire. C’est exactement l’inverse pour le potassium. En 1889, le chimiste allemand Walther Nernst fut le premier à formuler l’équation nous permettant de calculer le potentiel correspondant à l’équilibre de ces forces (autrement dit, le potentiel correspondant à l’arrêt de toute diffusion ionique de part et d’autre de la membrane). Au potentiel d'équilibre, le potentiel de membrane créé une force opposée égale au gradient chimique, si bien qu'aucune diffusion de l'ion étudié n'est possible.
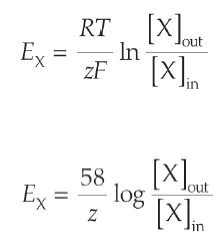 |
| Equation de Nernst |
Cette équation nous permet de calculer le potentiel d’équilibre du sodium (50mV) et du potassium (-90mV). On peut dès lors remarquer que le potentiel de repos de la membrane cellulaire, égal à -70mV, est assez éloigné du potentiel de repos du potassium et encore plus du sodium : spontanément, le sodium à tendance à vouloir entrer dans la cellule et le potassium à en sortir ! Heureusement, la membrane cellulaire et ses protéines veillent au grain, et repoussent toute velléité de passage !
Tout cela fonctionne bien chez un neurone au repos. Son potentiel de membrane est activement maintenu à -70mV, et chacun reste du bon côté de la membrane. Cependant, tout cela change drastiquement lorsqu’on le stimule.
La stimulation d’un neurone entraîne l’ouverture, progressive, de certains canaux membranaire perméables au sodium : aussitôt, des ions s’engouffrent dans la cellule, la rendant un petit peu moins négative (son potentiel membranaire croît). Si cette stimulation est suffisante, jusqu’à atteindre un potentiel de membrane de -55mV, elle déclenche une dépolarisation brutale du neurone (c’est-à-dire un changement brutal de polarité : la cellule devient chargée positivement et le milieu extra-cellulaire négativement). Cette dépolarisation est permise par l’ouverture massive de canaux sodiques voltage-dépendant : le sodium, enfin libre de ses mouvements, se rue à l’intérieur de la cellule jusqu’à atteindre son potentiel de repos à lui, soit environs 50mV (en réalité, la dépolarisation d'arrête à 30mV car les canaux sodium sont inactivés).
La membrane n’a pas le temps de souffler que déjà, les canaux potassium s’ouvrent à leur tour ! Aussitôt, les ions potassium (si loin de leur potentiel de membrane propre, maintenant que celui de la membrane est à +50mV) quittent massivement la cellule, la rendant de plus en plus négative. Les concentration de potassium ne se stabiliseront qu’une fois leur potentiel de repos atteint, soit -90mV.
La cellule est alors hyper-polarisée : elle entre dans sa période réfractaire. Au cours de celle-ci, le potentiel de membrane très négatif empêche toute stimulation du neurone (à moins de le stimuler très fortement). C’est alors que se met en action la fameuse pompe Na/K ATPase, qui permettra de rétablir les concentrations ioniques de part et d’autre de la membrane.
C’est l’ensemble de ces étapes que l’on appelle un potentiel d’action, qui est donc la conséquence de flux ioniques de part et d’autre de la membrane cellulaire du neurone. Ce potentiel d’action se propage de proche en proche, la période réfractaire l’empêchant de « faire demi-tour ». C’est ce phénomène qui se cache derrière l’expression « un courant électrique se propage le long de l’axone » !
Il faut d’ores et déjà noter que les variations de concentrations ioniques sont infinitésimales lors de ce processus. Nous pouvons calculer grâce à l’équation de Nernst qu’elle n’excède pas quelques centièmes de pourcents de la concentration de base. Ainsi, les courants ioniques, s’ils ont un impact majeur sur la polarisation membranaire, ont un effet négligeable sur les gradients de concentration.
Ce processus de dépolarisation membranaire correspond à l’activation d’un neurone. Cette activation est le plus souvent la conséquence d’une libération de glutamate par le neurone pré-synaptique. Mais il existe dans le cerveau des neurones dits inhibiteurs, dont le neurotransmetteur principal est le GABA et dont le rôle n’est pas de déclencher un potentiel d’action, mais plutôt d’empêcher un tel phénomène en provoquant une hyperpolarisation du neurone.
Ces interneurones inhibiteurs induisent cette hyperpolarisation via des canaux potassium comme nous l’avons vu plus haut, mais aussi grâce à des canaux chlore. Tout comme le sodium, ses concentrations sont plus élevées en dehors de la cellule (103mmol/L versus 5mmol/L) mais à l’inverse de celui-ci, sa charge électrique est négative. L’équation de Nernst nous permet de calculer un potentiel d’équilibre à -80mV environs. Le chlore a donc tendance à vouloir rentrer dans la cellule pour atteindre son point d’équilibre, rendant le milieu intracellulaire d’autant plus négatif et donc aboutissant à une hyperpolarisation.
Lorsque le GABA (libéré par un interneurone inhibiteur) se lie à son récepteur, il déclenche une ouverture des canaux chlore permettant ainsi une hyperpolarisation du neurone post-synaptique. Le potentiel de membrane s’éloignant du seuil de -55mV, le déclenchement d’un potentiel de membrane devient plus difficile.
Ces deux processus (excitation et inhibition) existent de manière concomitante au sein d’un même neurone. En effet, chaque neurone forme plusieurs milliers de synapses avec les neurones alentours. Certaines sont activatrices (les synapses fonctionnant au glutamate typiquement) et certaines inhibitrices. L’ensemble de ces informations entrantes sont sommées au niveau du corps cellulaire du neurone. Si les influx excitateurs sont supérieurs aux inhibiteurs, un potentiel d’action sera créé au niveau de l’axone et se propagera vers de nouveaux neurones.
Le cerveau d’un fœtus fonctionne à l’envers ! [2, 10]
La balance entre le GABA inhibiteur et le glutamate excitateur est cruciale pour un fonctionnement cérébral efficient. De nombreuses pathologies, et en premier lieu l’épilepsie, sont causées en partie par un déséquilibre de ces fonctions. En contrepartie, certains traitement anti-convulsivants ont une puissante action activatrice des récepteurs GABA permettant de rétablir l’équilibre.
Cependant, cette harmonie est bien différente chez le cerveau fœtal. En effet, alors que le bébé est encore dans la ventre de sa mère, le GABA cérébral n’est pas inhibiteur… Mais excitateur ! Comment expliquer ce revirement de situation ? Quelle est son utilité ? A quel moment bascule-t-on d’un fonctionnement fœtal à un fonctionnement adulte ?
Le cerveau d’un fœtus ne peut pas être considéré comme un cerveau adulte miniature. Il est le siège de processus développementaux majeurs, dont beaucoup perdureront jusqu’à l’adolescence.
A la fin des années 1980, une équipe de chercheurs français a fait une découverte étrange : l’activation des récepteurs GABA à la membrane de neurones hippocampiques de souris n’entraînait pas d’hyperpolarisation comme attendu, mais au contraire déclenchait un potentiel d’action.
Comment expliquer que le GABA, inhibiteur chez l’adulte, soit excitateur chez le fœtus ? Nous l’avons vu, la dépolarisation d’un neurone survient lorsqu’on augmente son potentiel de membrane, initialement à -70mV, jusqu’à un seuil de -55mV qui permet un renversement brutal de la polarité (qui devient positive). Chez l’adulte, cela est permis notamment par l’ion sodium dont le potentiel d’équilibre est de +60mV. Au contraire, l’ouverture des canaux chlore (ion dont le potentiel d’équilibre est de -80mV) permet de s’éloigner du seuil fatidique et de rendre moins probable l’apparition d’un potentiel d’action.
C’est justement au niveau du chlore que se trouve la réponse de notre énigme. A l’âge adulte, sa concentration intracellulaire est égale à 5mmol/L envions. Or, elle est légèrement plus importante chez le fœtus, de l’ordre de 20mmol/L. Cette différence a une conséquence majeure : si nous reprenons l’équation de Nernst, nous calculons un potentiel d’équilibre qui n’est plus de -80mV, mais de -40mV, qui se trouve donc au dessus du seuil de dépolarisation ! C’est ainsi que la stimulation GABAergique provoquera la sortie (et non l’entrée comme chez l’adulte) d’ions chlore et pourra aboutir à l’apparition d’un potentiel d’action !
Cette fonction activatrice du GABA n’est pas anodine. En effet, la maturation du système GABAergique est plus précoce que celle du système glutamatergique. Ainsi donc, aux premiers stades de développement, l’excitation neuronale repose exclusivement sur le GABA. Cela a son importance car c’est l’activité neuronale qui va moduler l’architecture de l’ensemble des réseaux cérébraux ! L’administration de drogues ou de médicaments qui interfèrent dans le fonctionnement du GABA (comme la Dépakine) peut avoir des conséquences désastreuses sur le cerveau en construction, aboutissant à un risque majeur de trouble psychiatrique (troubles du spectre de l’autisme par exemple) ou neurologique (épilepsie).
Ainsi donc, la mise en place des circuits cérébraux (dépendante de l’activité neuronale) repose exclusivement sur le GABA aux stades précoces du développement. Les synapses glutamatergiques, qui nécessitent un neurone plus mature pour se mettre en place, sont formées dans un second temps. Alors que les réseaux GABAergiques et glutamatergiques se complexifient progressivement, un besoin d’inhibition devient nécessaire afin d’harmoniser le fonctionnement de ces ensembles de neurones. C’est à ce moment là que l’action du GABA s’inverse, au cours d’un processus fortement influencé par l’activité neuronale et donc par l’environnement. Le switch semble en effet bloqué si l’on injecte aux souris une drogue inhibant l’action du GABA (cela ne semble pas être le cas pour le glutamate). Ce serait donc le GABA lui même qui induirait (en partie) son propre switch fonctionnel !
Les explications du développement séquentiel des systèmes GABA et glutamatergiques sont peu claires, et les explications hypothétiques. Nous pouvons cependant imaginer qu’une formation en parallèle de ces 2 systèmes, en respectant le fonctionnement adulte (GABA inhibiteur), serait d’une complexité incroyable. En effet, cela impliquerait la formation parfaitement synchronisée et en tout point et à chaque moment de la balance inhibition/excitation, dans un processus extrêmement rigide et peu sensible à l’environnement. Cela ne correspond pas à ce que l’on observe. Au lieu de cela, la maturation précoce du système GABAergique permet une excitation douce et prolongé des neurones immatures, adaptée au développement des circuits neuronaux et permettant, dans un second temps, la formation du système glutamatergique.
Mais comment expliquer que le GABA devienne inhibiteur après la naissance ? La membrane neuronale abrite de nombreuses protéines et en particulier des pompes ioniques. Nous avons déjà évoqué la pompe Na/K ATPase, qui permet de chasser le sodium de la cellule et d’absorber du potassium. Il existe aussi plusieurs types de pompes à chlore, et en particulier deux protéines nommées NKCC1 et KCC2, qui ont des fonctions opposées.
 |
| Neurone immature. NKCC1 est plus exprimé que KCC2 (qui mature plus tardivement), entraînant une entrée de chlore dans le neurone et une action excitatrice du GABA. |
NKCC1, qui est capable de transporter des ions chlore, sodium et
potassium de l’extérieur vers l’intérieur de la cellule, est
fonctionnel très tôt dans le développement cérébral, au
contraire de KCC2 (dont la fonction est au contraire de chasser le
chlore de la cellule). Ainsi donc, l'influence de NKCC1 sera très
importante au cours de la vie fœtale, expliquant la majoration de la
concentration intracellulaire de chlore. Petit à petit, la
maturation plus tardive de KCC2 permettra de vider la cellule de son chlore,
d’abaisser le potentiel d’équilibre de l’ion et de basculer
vers une action inhibitrice du GABA.
 |
| Neurone mature. KCC2 est pleinement exprimé, permettant une sortie du chlore intracellulaire et le développement de la fonction inhibitrice. |
Il existerait une certaine hétérogénéité dans la maturation fonctionnelle du système GABAergique. Ainsi, à la naissance, certains interneurones seraient matures (inhibiteurs) et d’autres seraient toujours immatures (donc excitateurs). Il semble cependant que la fin de ce processus soit assez précoce, au cours des toutes premières semaines de vie. De manière générale, le cerveau du nouveau-né est toujours en construction, et il le restera encore pour plusieurs années !
Il faut cependant, comme devant tout domaine de recherche et toute étude, rester extrêmement prudent. Les données dont nous parlons sont séduisantes, mais elles sont imparfaites. Tout d’abord, elles proviennent pour la plupart d’études chez la souris, voire in vitro. Leur extrapolation à l’Homme doit donc se faire très prudemment. Ces études se heurtent de plus à des limitations méthodologiques [11]. En effet, les méthodes de détection du canal NKCC1 sont imparfaites. Ce dernier existe sous plusieurs versions, que l’on appelle isoformes (A et B). Or, les anticorps utilisés dans nos études détectent très mal l’isoforme B, qui est pourtant la version de NKCC1 prépondérante dans le cerveau. Le profil développemental que l’on observe pourrait donc être faussé, étant donné qu’il ne représente que celui de l’isoforme A.
Il faut aussi prendre en compte une possible contamination des autres cellules cérébrales. La protéine NKCC1 est en effet exprimée aussi bien dans les cellules gliales que dans la paroi des vaisseaux sanguins. Le taux que l’on mesure pourrait donc être faussé par cette origine extra-neuronale.
Enfin, une proportion importante d’études portent sur l’hippocampe. Bien qu’il existe des arguments pour que les variations développementales observées soient présentes dans l’ensemble du cortex, les données pour ce dernier sont moins robustes.
Il faut donc rester prudent dans l’interprétation que nous pouvons faire de ces études. Il s’agit de théories séduisantes mais elles représentent un champs de recherche en cours avec son lot d’incertitudes. De futures études sont nécessaires, et certaines contrediront probablement certaines données.
Ocytocine, aussitôt fait ! [4]
Chez la souris, la bascule inhibitrice du GABA semble se faire extrêmement rapidement, en quelques heures à peine, lors de la naissance du petit souriceau. La chute brutale du chlore intracellulaire se retrouve dans tout le cortex, et nous incite donc à trouver une origine globale à ce changement brutal.
L’accouchement, nous l’avons vu, est un moment critique qui est déclenché par une sécrétion massive et brutale d’ocytocine par le cerveau maternel. Cette hormone agit sur l’utérus et déclenche les contractions. Serait-il possible qu'elle soit à l’origine du switch GABAergique, dans une danse cérébrale entre mère et bébé ?
Pour le savoir, des chercheurs ont (toujours chez la souris) administré in vitro un antagoniste de l’ocytocine (c’est-à-dire une molécule capable d’inhiber son action). Cela entraînait une stabilité notoire des concentrations intracellulaires de chlore, et donc le maintient des capacités excitatrices du GABA ! De plus, l’ocytocine semble bien provenir du cerveau maternel car le cerveau fœtal n’en produit pas !
Ce signal est d’autant plus important qu’il permettrait, selon certains chercheurs, de protéger le cerveau du bébé au cours de l’accouchement. En effet, cette « mise au repos » des neurones permettrait de les rendre plus résistants au manque d’oxygène inévitable lors de l’accouchement. Il s’agit là d’une explication particulièrement séduisante : ce serait le même signal qui déclencherait l’accouchement, qui préviendrait le bébé de l’imminence de sa naissance et qui protégerait son cerveau tout au long de ce périple tumultueux !
Cependant, aussi séduisante soit-elle, il est difficile d’extrapoler cette explication à l’Homme. En effet, les données indiquent une diminution du chlore intracellulaire plus précoce chez nous, au cours du dernier trimestre de la grossesse. Alors que le GABA reste partiellement excitateur plusieurs semaines après la naissance chez le rat, il semble déjà avoir viré de bord chez le fœtus humain. Nous ne savons pas bien à l’heure actuelle comment et par quels mécanismes se fait ce switch chez l’Homme. Il est possible que l’ocytocine entre en jeu, mais les mécanismes sous-jacents sont sans doute plus complexes.
L’ocytocine représente pourtant un espoir thérapeutique important. L’hormone pourrait être utilisée comme un neuroprotecteur puissant chez le nouveau-né. Mais cet effet semble plus lié à une modulation immunitaire ou hormonale (en particulier en diminuant le taux de glucocorticoïdes dans la sang) qu'à une action directe sur les neurones.
En conclusion, le cerveau du bébé est un organe incroyable, dont la malléabilité extrême le rend plus complexe encore que le cerveau adulte. Son fonctionnement est unique, et les contraintes sont telles que cela peut aboutir à un changement total de paradigme et de profil d’activation pour un même neurotransmetteur ! La naissance s’accompagne de changements majeurs dans le cerveau du bébé, comme nous l’indique les études de modèles animaux. Cependant, il reste difficile à l’heure actuelle d’extrapoler ces données à l’Homme.
SOURCES :
- [1] : Ami, Olivier, et al. "Three-dimensional magnetic resonance imaging of fetal head molding and brain shape changes during the second stage of labor." Plos one 14.5 (2019): e0215721.
- [2] : Ben-Ari, Yehezkel. "Excitatory actions of gaba during development: the nature of the nurture." Nature Reviews Neuroscience 3.9 (2002): 728-739.
- [3] : Watanabe, Miho, and Atsuo Fukuda. "Development and regulation of chloride homeostasis in the central nervous system." Frontiers in cellular neuroscience 9 (2015): 371.
- [4] : Zinni, Manuela, et al. "Modulating the oxytocin system during the perinatal period: a new strategy for neuroprotection of the immature brain?." Frontiers in neurology 9 (2018): 229.
- [5] : Ben-Ari, Yehezkel. "Is birth a critical period in the pathogenesis of autism spectrum disorders?." Nature Reviews Neuroscience 16.8 (2015): 498-505.
- [8] : https://www.fun-mooc.fr/c4x/amu/38005S02/asset/Ch5Cours-Transmission_synaptique_GABA.pdf
- [9] : https://fr.wikipedia.org/wiki/Potentiel_de_repos
- [10] : http://ipubli-inserm.inist.fr/bitstream/handle/10608/6235/MS_2007_8-9_751.html#R1
- [11] : Kharod, Shivani C., Seok Kyu Kang, and Shilpa D. Kadam. "Off-label use of bumetanide for brain disorders: an overview." Frontiers in neuroscience 13 (2019): 310.
CREDITS IMAGES ET VIDEOS :
- https://www.youtube.com/watch?v=oK3esXMQxaI
- https://www.youtube.com/watch?v=3t5ebbxFRrI
- Image par OpenClipart-Vectors de Pixabay
- Image par Bruno /Germany de Pixabay
- Image par Clker-Free-Vector-Images de Pixabay